| Пояснительный текст |
Данный файл включает [ таблицы ] в формате Exel: на первом листе помещен перечень ассоциаций доминантной классификации для лесов южной тайги и подтайги (хвойно-широколиственных лесов) в том виде, в каком эти единицы приведены в литературе; второй лист содержит литературные источники; на третьем листе помещен редуцированный список ассоциаций на основе информации из первого листа. Таблицы существуют в виде архива и открываются в отдельном окне при нажатии на выделенное в тексте слово.
Перечень ассоциаций расположен в той последовательности, которая взята за основу на данном сайте. В пределах подсекций ассоциации (типы леса) сгруппированы по доминантам древесного полога (группы типов леса = группы ассоциаций). Поскольку многие авторы ощущали необходимость более мелкого группирования полученных единиц, мы дополнили таблицы колонкой под названием «Подгруппы, ряды, циклы», т.к. эти названия употребляются разными авторами в сходном объеме. Поскольку только в отдельных случаях было осуществлено подобное деление (основанное на ведущих доминантах травяно-кустарничкового или мохового яруса), мы посчитали возможным продлить эту работу. Наши предложения в соответствующей колонке приведены нежирным шрифтом. В этой колонке жирный шрифт соответствует названиям, которые предложены авторами.
Названия ассоциаций приведены на русском и по-латыни в том случае, если они даны у соответствующих авторов; если латинское название у автора(ов) отсутствует, ячейка остается пустой.
|
| Обсуждение некоторых проблем и замечания, возникшие во время этой работы
|
1. Поскольку доминантная система построена на объединении сообществ со сходными доминантами, а последние характеризуются часто широким экологическим диапазоном, ориентация на доминанты травяного яруса приводит к тому, что сообщества со сходными доминантами могут существенно отличаться по своему флористическому составу. Особенно хорошо это видно на примере таких видов как Oxalis acetosella и Filipendula ulmaria. Стоит отметить, что ельники кисличные довольно разнообразны по составу, а то, что не существует такого единого типа леса, отмечала еще Е.Л. Любимова (1957). Кислица – это вид, который одним из первых занимает нарушенные участки почвы и становиться доминантом при отсутствии конкурентов и тем самым часто маркирует предшествующие нарушения.
|
2. По этой причине необходимо не только выявлять доминанты, но и обращать внимание на видовой состав и соотношение эколого-ценотических групп. Ошибок можно избежать, если последовательно решать вопрос о положении сообщества в иерархии единиц: сначала найти секцию, затем подсекцию, потом группу и подгруппу, где помещено данное сообщество.
|
3. Необходимо учитывать, что практически все типы леса (ассоциации) имеют переходные состояния и предпочтительнее не выделять их в особые единицы, а рассматривать как варианты близлежащих ассоциаций. Кроме того, практически каждая локальная выборка имеет свои виды, отсутствующие в другой выборке из этой же зоны. Здесь также стоит заметить, что отнесение того или иного сообщества к определенной классификационной единице требует анализа целой выборки (от 7 до 10 и более описаний), а не единственного описания. Сокращенные перечени видов, которые приводятся только в виде текста, очень часто затрудняют отнесение сообществ к той или иной единице из-за недостаточности информации.
|
4. Некоторые весьма физиономичные виды (чаще всего - крупные папоротники), привлекая внимание исследователя, способствуют выделению особых крупно-папоротниковых ассоциаций. Анализ сообществ в широком спектре условий приводит к выводу, что это не какой-то определенный тип, а совокупность сообществ разной экологии и разного положения в классификационной системе. Стоит отметить, что такую особенность папоротниковых типов подчеркивал еще А.А.Корчагин (1940). По этой причине имеет смысл выявлять ведущую эколого-ценотическую группу помимо папоротников.
|
5. Как видно из списка ассоциаций, в рамках каждой подсекции они оказываются неоднородными по нескольким направлениям. Во-первых, внутри секции они группируются по видам-доминантам или содоминантам, что может соответствовать как изменению микроусловий (малый градиент факторов), так и сукцессионному состоянию сообществ. Во-вторых, они отличаются набором диагностических видов при сходстве видов-доминантов, что обычно соответствует или смене мезоусловий (средний градиент факторов) или разному географическому положению сообществ.
|
6. Для большинства секций и подсекций действует правило однотипности соотношений в структуре сообществ. Так, большинство лесов с преобладанием бореальных кустарничков, как правило, имеют развитый моховой покров; однако встречаются сообщества, в которых эта связь нарушается. Такое нарушение соотношений встречается практически во всех подсекциях. Эти случаи отмечены в двух первых столбцах красным цветом и предполагаемые причины отклонения прописаны в Примечаниях. Возможно, что одной из причин таких отклонений являются какие-то предшествующие нарушения и нередко - создание культур. |
7. Наиболее сложно организована подсекция в пределах травяной секции, которая соответственно имеет название «сложной». Здесь мы можем наблюдать большое разнообразие сочетаний между составом и структурой разных ярусов сообществ. Обозначим наиболее важные из них.
А. Древесный ярус монодоминантный (ель, сосна), травяной ярус – полидоминантный и включает как бореальные, так и неморальные виды в близких пропорциях; моховой покров не развит или развит слабо.
Б. Древесный ярус монодоминантный (ель, сосна), травяной ярус преимущественно неморальный; моховой покров не развит или развит слабо. Такие леса мы помещали в секцию неморальных.
В. Древесный ярус поли-олигодоминантный (ель+широколиственные; сосна+широколиственные); травяной ярус – полидоминантный и включает как бореальные, так и неморальные виды в близких пропорциях; моховой покров не развит или развит слабо. Это наиболее распространенный вариант гемибореальной лесной растительности в южной тайге и подтайге Европейской России.
Г. Древесный ярус монодоминантный (ель), в травяном ярусе преобладают неморальные виды; моховой покров из бореальных видов развит хорошо (встречается на Южном Урале: Горчаковский, 1956).
В сложных типах сообществ монодоминантность верхнего яруса предположительно рассматривается как отражение хозяйственной деятельности (пожары, вырубка и создание культур, прочистки, формирование залежей и их последующее заселение). К сожалению, не удалось встретить каких-либо гипотез относительно значительного развития (до 80%) мохового покрова в лесах сложной подсекции. |
8. Для каждой ассоциации мы стремились (на сколько это было возможно) найти соответствующий ей синтаксон на уровне ассоциации в эколого-флористической классификации. Для этого использовались, как группы диагностических видов, так и по возможности весь список видов, если подобная информация была доступна. Естественно, что в ряде случаев эта дефиниция была под сомнением, такая ситуация отмечена вопросительным знаком. Если для синтаксона имелись синонимы, они также приводились. Поскольку полные названия ассоциаций есть в соответствующих таблицах сайта (см. продромус для соответствующей зоны), в приводимых таблицах названия синтаксонов даны в краткой форме. |
9. Достаточно часто оказывалось, что соответствующий синтаксон нам не удалось встретить в доступной литературе; в этом случае делалась отметка «не указан». Подобные случаи встречаются не столько редко, что свидетельствует о необходимости проработки соответствующих сообществ с позиций эколого-флористической классификации на более обширном геоботаническом материале. |
10. Очень неудачным в названиях ассоциаций является прилагательное «разнотравные», которое часто используется исследователями. Разнотравье в каждой секции очень разное по составу, и по такому названию не понятно, что именно автор имеет ввиду; положение таких сообществ его каждый раз приходится уточнять по списку видов. |
| Проведенная работа показывает, что насущной задачей становиться отработка иерархии единиц и унификация русских и латинских названий единиц всех иерархий в доминантной классификации. Только в этом случае можно требовать от исследователей строго придерживаться правил Кодекса, проект которого опубликован в 2001 г. (Нешатаев, 2001). |
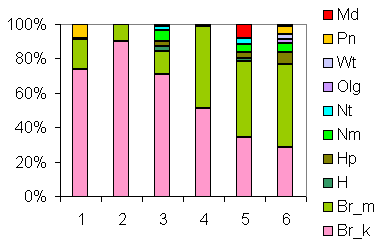
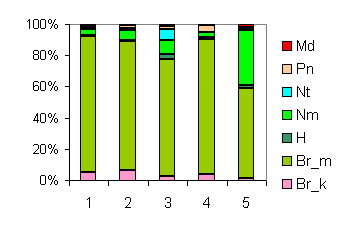
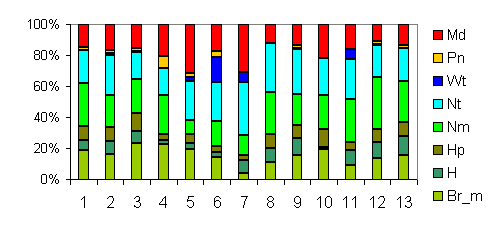
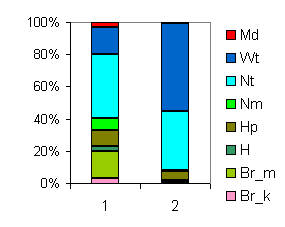
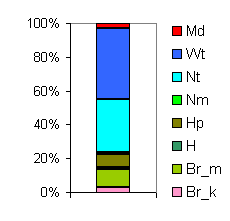
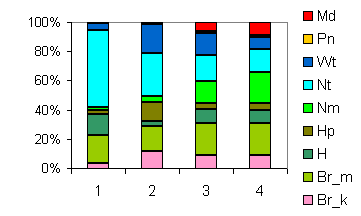
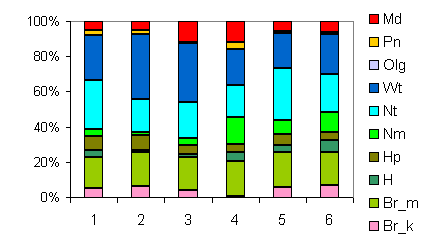
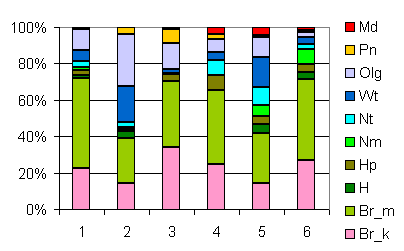
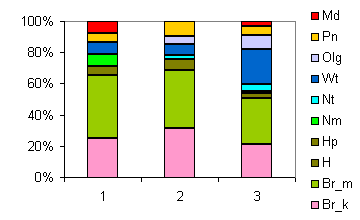
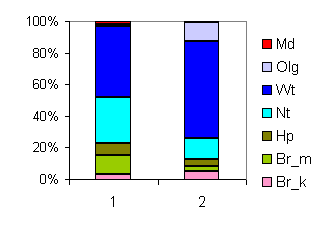
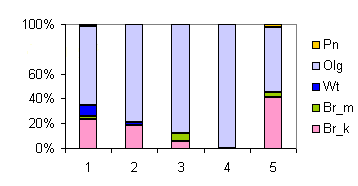
| Таблица спектров отдельных ассоциаций |
| Секция |
Подсекция |
Доминанты |
Содоминанты и дополнительные виды |
Доминанты яруса С в разных сочетаниях |
Спектр эколого-ценотических групп |
| Лишайниковая секция |
Лишайниковая подсекция |
Сосна |
|
Calluna vulgaris, Festuca ovina, Koeleria glauca, Chimaphilla umbellata, Vaccinium vitis idaea |
3ll01.gif |
| Лишайниковая секция |
Зеленомошно-лишайниковая |
Сосна |
береза |
Calluna vulgaris,Vaccinium vitis idaea, Calamagrostis epigeios, C. arundinaceae |
3lz01.gif |
| Зеленомошная |
Мезоксерофильная |
Сосна |
береза |
Festuca ovina, Avenella flexuosa, Koeleria glauca, K. grandis, Calamagrostis epigeios |
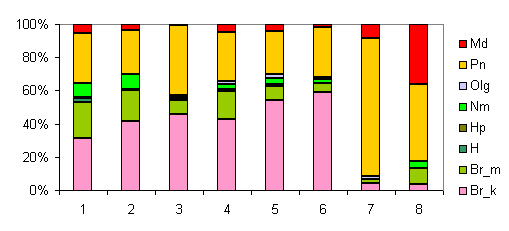
3zk01.gif |
| Зеленомошная |
Кустарничковая |
Сосна |
ель // дуб |
Calluna vulgaris, Vaccinium vitis idaea, Calamagrostis arundinaceae |
3zk01.gif |
| Зеленомошная |
Кустарничковая |
Сосна |
ель, береза, осина // дуб, липа |
Vaccinium mytrillus, V. vitis idaea, Calamagrostis arundinaceae, Pteridium aquilinum |
3zk01_1.gif |
| Зеленомошная |
Кустарничковая |
Ель (Picea abies, Picea obovata) |
сосна, береза, осина //дуб, липа |
Vaccinium mytrillus, V. vitis idaea, Calamagrostis arundinaceae, Pteridium aquilinum |
3zk04.gif |
| Зеленомошная |
Мелкотравная |
Ель (Picea abies, Picea obovata) |
сосна, береза, осина, ольха серая // дуб, липа |
Maianthemum bifolium, Oxalis acetosella,Gymnocarpium dryopteris, Equisetum sylvaticum, Vaccinium mytrillus, Calamagrostis arundinaceae |
3zm04.gif |
| Зеленомошная |
Мелкотравная |
Ель (Picea abies) |
береза // дуб, липа |
Maianthemum bifolium,Oxalis acetosella,Gymnocarpium dryopteris, Rubus saxatilis, Calamagrostis arundinacea |
3hm04_1.gif |
| Зеленомошная |
Мелкотравная |
Сосна |
ель, береза // дуб |
Oxalis acetosella, Rubus saxatilis,Vaccinium mytrillus, Calamagrostis arundinaceae |
3zm01.gif |
| Травяная |
Мелкотравная
(неморально-бореальная) |
Ель |
сосна, береза пушистая,осина, серая ольха // липа |
Oxalis acetosella, Rubus saxatilis, Equisetum sylvaticum, Galeobdolon luteum |
3hm04.gif |
| Травяная |
Мелкотравная
(неморально-бореальная) |
Береза повислая, пушистая |
|
Oxalis acetosella, Calamagrostis arundinaceae, Rubus saxatilis |
3hnb06.gif |
| Травяная |
Сложная (бореально-неморальная) |
Ель (Picea abies) |
сосна // дуб, липа |
состав полидоминантный - Oxalis acetosella, Equisetum sylvaticum, Vaccinium mytrillus, Galeobdolon luteum, Aegopodium podagraria, Carex pilosa, Calamagrostis arundinaceae |
3hbn04_w.gif |
| Травяная |
Сложная (бореально-неморальная) |
Ель (Picea abies) |
липа, дуб, клен, вяз // береза, осина |
состав полидоминантный: Oxalis acetosella,Galeobdolon luteum, Aegopodium podagraria, Carex pilosa |
нет |
| Травяная |
Сложная (бореально-неморальная) |
Осина |
береза // ель, дуб, ольха серая |
Rubus saxatilis,Oxalis acetosella, Aegopodium podagraria, Pteridium aqilinum |
3hbn06_p.gif |
| Травяная |
Сложная (бореально-неморальная) |
Сосна |
липа, дуб //ель, береза, осина |
состав полидоминантный - Oxalis acetosella, Vaccinium mytrillus, Galeobdolon luteum, Carex pilosa, Mercurialis perennis, Calamagrostis arundinaceae |
3hnb01.gif |
| Травяная |
Высокотравная |
Ель (Picea obovata, Picea abies) |
липа, пихта // береза, осина |
полидоминантный - Aconitum septentrionale, Aegopodium podagraria, Oxalis acetosella, Maianthemum bifolium, Filipendula ulmaria, Geum rivale |
3hh04.gif |
| Травяная |
Высокотравная |
Ель (Picea obovata) |
липа, пихта // клен, вяз, береза |
Aegopodium podagraria, локально - Aconitum septentrionale, Urtica dioica |
нет |
| Травяная |
Высокотравная |
Береза (оба вида) |
// липа, пихта, ель |
Aegopodium podagraria, Aconitum septentrionale, Cicerbita uralensis, Crepis sibirica, Urtica dioica, Filipendula ulmaria |
3hbn06.gif |
| Травяная |
Высокотравная |
Осина |
// береза, сосна, ель |
Aegopodium podagraria, Aconitum septentrionale, Cicerbita uralensis, Crepis sibirica, Urtica dioica, Filipendula ulmaria |
нет |
| Травяная |
Неморальная |
Липа/дуб |
// клен остролистный , вяз, ясень, береза, осина |
Galeobdolon luteum, Aegopodium podagraria, Carex pilosa, Mercurialis perennis |
3ne10.gif |
| Травяная |
Неморальная |
Липа/дуб |
ель// клен остролистный, вяз, ясень, береза, осина |
Galeobdolon luteum, Aegopodium podagraria, Carex pilosa, Mercurialis perennis |
нет |
| Травяная |
Неморальная |
Береза (оба вида) |
дуб, липа// клен остролистный, вяз, ясень, береза, осина |
Galeobdolon luteum, Aegopodium podagraria, Carex pilosa, Ajuga repens, Oxalis acetosella |
3ne06.gif |
| Травяная |
Неморальная |
Осина |
дуб, липа // клен остролистный, вяз, ясень, береза, осина |
Galeobdolon luteum, Aegopodium podagraria, Carex pilosa, Stellaria holostea, Ajuga repens |
нет |
| Травяная |
Нитрофильная |
Осина |
береза пушистая |
Geum rivale,Carex pilosa, Filipendula ulmaria,Crepis paludosa |
3hni06.gif |
| Травяная |
Нитрофильная |
Ольха серая |
//береза пушистая, ива козья, ель |
Urtica dioica,Filipendula ulmaria,Equisetum sylvaticum, E. pratense, Aegopodium podagraria, Oxalis acetosella, Matteuccia strutioptheris, Deschampsia cespitosa |
3ni_13.gif |
| Травяная |
Нитрофильная |
Ольха черная |
липа, ель, береза пушистая |
Urtica dioica,Filipendula ulmaria, Aegopodium podagraria, Matheuccia strutioptheris, Athyrium filix- femina |
3ni07.gif |
| Болотно-травяная |
Эвтрофно-болотная |
Ольха черная |
береза пушистая |
Filipendula ulmaria, Phragmithes australis, Calamagrostis canescens, Calla palustris, Athyrium filix-femina; во втором подъярусе - Aegopodium podagraria, Geum rivale |
3wf07.gif |
| Болотно-травяная |
Эвтрофно-болотная |
Ольха черная |
береза// ель, осина |
Filipendula ulmaria, Carex gracilis, C. cespitosa, C. vesicaria, Galium palustre, Glyceria aquatica |
нет |
| Болотно-травяная |
Эвтрофно-болотная |
Береза пушистая |
ольха черная |
Filipendula ulmaria, Phragmithes australis, Calamagrostis canescens, Calla palustris, Athyrium filix-femina; во втором подъярусе - Aegopodium podagraria, Geum rivale |
3wf06.gif |
| Болотно-травяная |
Эвтрофно-болотная |
Ольха серая |
береза пушистая |
Phragmithes australis, Calamagrostis canescens, Carex cespitosa |
нет |
| Болотно-травяная |
Мезотрофно-болотная |
Ель |
береза пушистая, ольха черная |
Filipendula ulmaria, Calamagrostis canescens (на востоке C. purpurea), Carex cespitosa, Galium palustre |
3hg04.gif |
| Болотно-травяная |
Мезотрофно-болотная |
Береза пушистая |
ель, ольха черная |
Filipendula ulmaria, Calamagrostis canescens (навостоке C. purpurea), Carex cespitosa |
3hg06.gif |
| Сфагновая |
Долгомошно-сфагновая |
Сосна |
береза пушистая, ель |
Molinia caerulea, Vaccinium myrtillus |
3sm01.gif |
| Сфагновая |
Долгомошно-сфагновая |
Ель |
береза пушистая, сосна |
Vaccinium myrtillus, V. vitis idaeaEquisetum sylvaticum, Carex globularis |
3sm04t.gif |
| Сфагновая |
Долгомошно-сфагновая |
Береза пушистая |
ель, сосна, осина |
Vaccinium myrtillus, V. vitis idaea, Molinia caerulea, Carex globularis, Vaccinium uliginosum |
3sm06.gif |
| Сфагновая |
Травяно-сфагновая |
Береза пушистая |
ель, сосна |
Carex canescens, Carex lasiocarpa, Carex elongata, Calamagrostis canescens (навостоке C. purpurea), Galium palustre |
3sg06t.gif |
| Сфагновая |
Травяно-сфагновая |
Ольха черная |
ель, береза пушистая |
Calamagrostis canescens (навостоке C. purpurea), Equisetum fluviatile, Phragmithes australis, Menyanthes trifoliata,Calla palustris |
3sg07.gif |
| Сфагновая |
Кустарничково-сфагновая |
Сосна |
береза пушистая, береза повислая |
Chamaedaphne calyculata, Andromeda polifolia, Eriophorum vaginatum, Oxycoccus palustris |
3sk01.gif |
|
Дополнительная литература |
Любимова Е.Л. Очерк растительности природных районов Московской области // Материалы по физической географии СССР. Тр. Ин-та географии АН СССР. Т. 71. № 2. 1957. С.42-82 .
Нешатаев В.Ю. Проект Всероссийского кодекса фитоценологической номенклатуры//
Растительность России\. СПб. № 1. 2001. С. 57-69. |
|

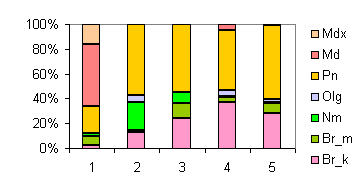{kind=link}
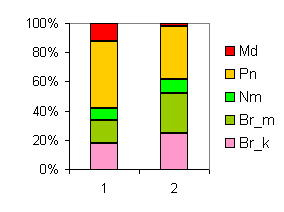{kind=link}
{kind=link}
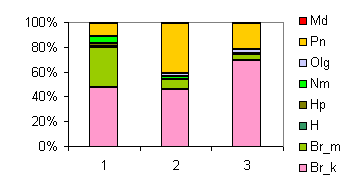{kind=link}
{kind=link}
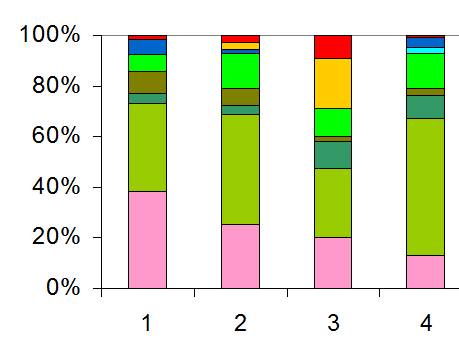{kind=link}
{kind=link}
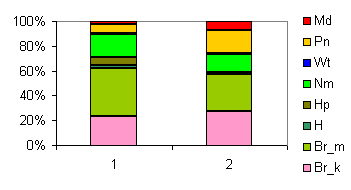{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
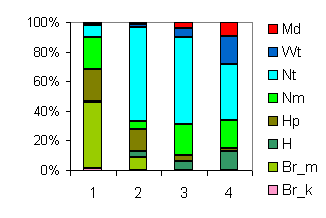{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
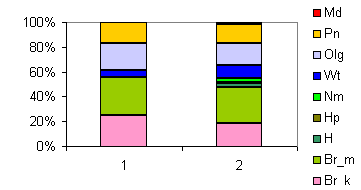{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}