Типология |
| Группа типов: |
| ельники сложные
(бореально-неморальные) |
| Piceeta
composita (boreo-nemoherbosa) |
| Зональное положение: южная тайга
и подтайга |
| Класс: гемибореальные восточноевропейские
хвойно-широколиственные леса |
| Секция: травяная herbosa
|
| Подсекция: сложные (бореально-неморальные)
composita (boreo-nemoroherbosa) |
| |
| Синтаксономическое положение |
| Эколого-флористическая классификация |
| Эта группа сообществ объединяет
леса с одновременным присутствием как бореальных, так
и неморальных видов, но участие неморальных
обычно выше по сравнению с бореальными. На этом основании
данные сообщества относят к классу Querco-Fagetea
Br.-Bl. et Vlieger in Vlieger 1937 em. Klika
1939, а в пределах класса – к порядку Fagetalia
sylvaticae Pawl. in Pawl., Sokol. et Wallish. 1928 и союзу
Querco-Tilion Solomesc et Laivinsh in Solomesc 1993. Эта группа типов соответствует асс. Rhodobryo
rosei-Piceetum abietis Korotkov
1986, которая характерна для южной тайги и подтайги на западе
и в центре Русской равнины, а на востоке достигает Костромской
области.
В Татарии (Порфирьев, 1950) происходит постепенная смена на другую ассоциацию (ранее - Tilio-Piceetum obovatae Schubert,Jager et Mahn 1979 ex.Mirkin et Solometch 1990, позднее она вошла в состав асс. Chrysosplenio alternifolii-Piceetum obovatae Martynenko et Zhigunova 2007 ), которая обогащена высокотравными видами урало-сибирского ареала. |
| Доминантная классификация |
Наиболее часто встречающееся
название этой группы – сложные ельники, или Piceeta
composita (Рысин,
Савельева, 2002); оно в большинстве случаев используется
для более западных вариантов бореально-неморальных ельников.
У разных авторов можно встретить довольно разнообразный
набор названий: Piceetum tiliosum
(Соколов, 1931), Abiegno-Piceetum oxalidoso-nemorosum,
Piceetum oxalidoso-nemorosum
(Порфирьев, 1950), а также еловые
леса и липо-ельники южной тайги и подтайги с разными
доминантами травяного яруса – зеленчуком (Galeobdolon
luteum), осокой волосистой (Carex pilosa),
снытью (Aegopodium podagraria), кислицей (Oxalis
acetosella) (Ниценко, 1961; Яруткин, 1981; Речан
и др., 1993; Савельева, 2000; Рысин, Савельева, 2004;
Василевич, 2004).
Поскольку сложные ельники и хвойно-широколиственные леса,
несмотря на разное соотношение в древостое ели и широколиственных
видов, демонстрируют высокую степень сходства по общему
флористическому составу, мы приводим здесь описание тех
и других совместно. |
Фитоценотическая характеристика |
Древесный ярус (А):
Сомкнутость может варьировать в широких пределах (0,3-0,8),
что зависит в значительной степени от возрастной структуры
древостоя. В его составе преобладают темнохвойные виды:
ель (Picea abies – на западе, или ее гибриды с Picea obovata в сочетании с пихтой (Abies sibirica)
на востоке. Обычно в качестве дополняющих видов небольшое
участие принимают мелколиственные – береза (Betula
pubescens, B. pendula), осина (Populus tremula),
ольха серая (Alnus incana) и широколиственные
– преимущественно липа (Tilia cordata), а в южной
части ареала группы - дуб (Quercus robur). Соотношение
видов в древесном ярусе может варьировать в широких пределах
даже на ограниченной территории: от преобладания ели до
паритетного участия видов. Нередко формируется два подъяруса
(первый – высотой до 22-26 м; второй – до 14-16 м), при
этом широколиственные породы могут входить как в первый,
так и во второй подъярус. Например, весьма сложная структура
отмечена для хвойно-широколиственных лесов Чувашии (Яруткин,
1981): 1-й подярус 4Е3Б2Ос1Лп, 2-й подярус – 5Лп3Д2Е.
Ель распределена в сообществах неравномерно (Коротков,
Морозова, 1988): в результате воздействия на нее корневой
губки и большого черного усача образуются окна в пологе
древостоя. Участки под такими окнами часто зарастают кустарниками. |
| Ярус подлеска (B): Развит
в разной степени: от 30% (Коротков, 1991) до 60% (Шапошников
и др., 1988). В его состав входят лещина
(Corylus avellana), рябина (Sorbus aucuparia),
крушина (Frangula alnus), жимолость (Lonicera
xylosteum), шиповник (Rosa majalis), калина
(Viburnum opulus). Представлено возобновление
ели, березы, ольхи серой, липы, редко - дуба. Подрост
ели концентрируется в участках под окнами или в осиново-березовых
группировках (Яруткин, 1981). |
| Травяно-кустарничковый ярус
(C): Покрытие сильно варьирует в зависимости от развития
верхних ярусов сообщества, но чаще всего довольно высокое
– до 80-90%. Доминантами или содоминантами являются как
неморальные виды: сныть (Aegopodium podagraria),
осока волосистая (Carex pilosa), зеленчук желтый
(Galeobdolon luteum), так и бореальные: кислица
(Oxalis acetosella), майник (Maianthemum bifolium),
черника (Vaccinium myrtillus). На песчаных почвах
в состав содоминантов входит вейник лесной (Calamagrostis
arundinacea). Весной в этих лесах могут встречаться
эфемероиды – например, виды ветреницы (Anemonoides
ranunculoides, A. nemorosa). В окнах полога и на
вывалах (а также на небольших вырубках) мощно развиваются
виды крупных папоротников (Athyrium filix-femina,
Dryopteris filix-mas, D. expansa, D. carthusiana), формируя
крупнопапоротниковые варианты сообществ. |
Мохово-лишайниковый ярус
(D): Этот ярус развит слабо, хотя встречаемость отдельных
видов может быть высокой. Наиболее постоянны: Pleurozium
schreberi, Hylocomium splendens, Dicranum scoparium.
Весьма характерен Rhodobryum roseum, присутствуют
виды родов Plagiomnium и Brachythecium.
В Подмосковье, как северном (Речан и др., 1993 – стр.
46), так и в южном (Биогеоценотические исследования…,
1971 – стр. 9-35), встречается вариант сложных ельников
с моховым покровом (7-13%), который
рассматривается как признак культурфитоценоза, формирующегося
на старопахотных землях (Речан и др., 1993). Моховая синузия
наиболее успешно развивается в участках под сомкнутым
подростом ели, в черничных микрогруппировках и на нарушенных
участках. |
| Внеярусная растительность (E): Представлена
преимущественно эпифитными мхами (Коротков, 1991) и лишайниками
(Речан и др., 1993), а на востоке ареала группы – одревесневающей
лианой Atragene sibirica. |
| |
| Спектр эколого-фитоценотических
групп наземных ярусов: |
|
| Обозначения групп видов: Br_k
– бореальные кустарнички и вечнозеленые травы; Br_m
– бореальное мелкотравье; H – бореальное
высокотравье; Hp – крупнотравные папоротники;
Nm – неморальные травы; Nt
– нитрофильные травы; Wt – гигрофильные
травы; Olg – олиготрофно-гигрофильные
травы и кустарнички; Pn – мезоксерофильные
боровые травы; Md – луговые травы |
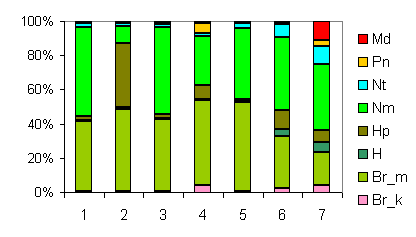
Ассоциация Rhodobryo-Piceetum в западном секторе зон:
1,2 - Северо-запад Европейской России (Василевич, 2004): Tilio-Piceetum*
(1 - неморальные, 2 - бореальные); 3,4 – там же: Corylo-Piceetum*;
5 – там же: Acero-Piceetum*;
6 – Тверская обл., Центрально-лесной заповедник (Шапошников
и др., 1988); в центральном секторе - 7 - Московская обл. (Коротков, Морозова,
1986).
*Авторские названия сообществ (Василевич, 2004). |
| |
|
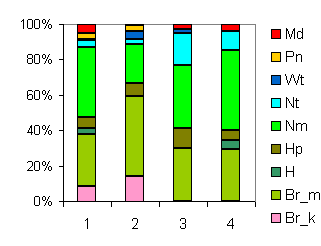
| Ассоциация Rhodobryo-Piceetum в восточном секторе зон представлена субассоциацией abietetosum sibiricae (Нижегородская обл.): 1,2 - Соколов, (1931): 1 - Piceetum tiliosum*, 2 - Piceetum oxalidosum*; 3 - неопубликованные описания А.И. Широкова ; 4 - Хомутова (1941). * Авторские названия сообществ. |
|
| Фотография сообщества
Фото 1:
Республика Марий Эл, нац. парк "Марий Чодра". Автор М.В.
Бекмансуров. Фото
2, Фото
3: Костромская обл., Кологривский р-н. Автор Д.Л.
Луговая. |
Характеристика разнообразия |
| Внутрисинтаксономическое
разнообразие |
Группа сложных ельников и
бореально-неморальных хвойно-широколиственных лесов весьма
разнообразна как по возрастной структуре древостоев, так
и по соотношению доминантов в разных ярусах. Древостой
может состоять как из одного поколения ели (Коротков,
1991), так и быть относительно или абсолютно разновозрастным
(Коренные южно-таежные леса, 1988; Широков, 1998). В восточной
части ареала группы в древостое принимает значительное
участие пихта - Abies sibirica (Турков, 1979,
1985; Смирнова и др., 2006).
В связи со сложной пространственной структурой и многовидовым
флористическим составом, включающим несколько потенциальных
видов-доминантов, в природе можно обнаружить большое число
различных сочетаний доминантов травяного покрова, что
и отражается в разнообразии названий отдельных типов (ельники
снытевые, волосистоосоковые, пролесниковые, зеленчуковые),
а также варианты с двумя-тремя содоминантами как в древесном
(Василевич, 2004), так и в травяно-кустарничковом ярусах.
В наиболее сохранившихся лесных массивах для этой группы
сообществ характерна абсолютно разновозрастная структура
древостоя (Коренные южно-таежные леса, 1988) и сложная
внутренняя организация с четко выраженной парцеллярной
мозаикой (Коротков, 1991; Широков, 1998; Кураева и др.,
1999). Для Центрально-лесного заповедника (Бобров и др.,1999)
показано, что распределение древесных видов в сложных
ельниках соотносится с неоднородностью оконной мозаики:
группы 100-летних елей сочетаются с ильмом (Ulmus
scabra); ель двух поколений - с разреженным ярусом
ильма и клена (Acer platanoides); относительно
недавно появившиеся окна заполнены 20-30-летним кленом
и ильмом; ель 230-240 лет сочетается с липой во втором
подъярусе. Для травяного покрова установлена связь с типами
западин микрорельефа: сухие западины с богатой почвой
характеризуются преобладанием неморальных видов; влажные
с богатой почвой – нитрофильными видами, влажные с бедной
почвой – заняты мхами (Polytrichum commune, видами
Sphagnum).
Ассоциация Rhodobryo-Piceetum
описана на территории Русской равнины и в ее составе выделено
три субассоциации (Заугольнова, Морозова, 2004). Субассоциация
caricetosum pilosae представлена
в центре Русской равнины (диагностируется
по Carex pilosa). Субассоциация typicum
(диагностируется по Hepatica nobilis, Anemonoides
nemorosa, Dryopteris expansa) первоначально
описана в западной части Русской равнины (Коротков, 1991)
. Однако анализ дополнительной литературы
показал, что в сложных ельниках Нижегородской области
(Соколов, 1931) тоже представлены некоторые диагностические
виды субассоциации typicum,
причем одновременно выражены и признаки третьей субассоциации,
типичной для восточной части ареала группы. Эта субассоциация
- abietetosum sibiricae -
встречается в Костромской и Нижегородской области и представляет
собой наиболее восточный вариант данной ассоциации.
Исследователи, использующие исключительно доминантную
классификацию, разделяют сложные ельники и липо-ельники
(Речан и др., 1993, стр. 34-35) на несколько более мелких
групп, названных «циклами» (волосистоосоковый, снытевый,
пролесниковый, кисличный). Эти «циклы» в ряде случаев
можно рассматривать как варианты ассоциации Rhodobryo-Piceetum,
различающиеся между собой не только доминантами травяного яруса, но и соотношением неморальных и бореальных видов. Такое
варьирование наблюдается на фоне сохранения общей структуры
эколого-фитоценотического спектра, которая свойственна
данной группе типов.
Л.П. Рысин (1961) расматривал варианты ельников с высоким
обилием кислицы в качестве особой ассоциации «ельник кислично-щитовниковый».
Здесь стоит заметить, что обилие кислицы характерно для
одного из этапов восстановительных сукцессий и, как показывает
сопоставление флористических списков и эколого-фитоценотической
структуры, ельники кисличные далеко не равнозначны: в
одних случаях в них преобладают бореальные виды (см. Ельники мелкотравно-зеленомошные), в других - присутствуют
на паритетных началах бореальные и неморальные виды (см.
Ельники мелкотравные
(неморально-бореальные)), в третьих – преобладают
неморальные виды (см. Ельники
неморальные). Неоднородность ельников-кисличников
как производных типов отмечал С.Ф. Курнаев (1968). |
| |
| Видовое разнообразие синтаксона
(по группам видов) - Открыть в отдельном окне. Число видов в выборках. Ассоциация Rhodobryo-Piceetum. |
| |
| Видовая насыщенность (среднее число видов на единицу
площади и диапазон). Сокращения: Rh-Pic car.pil - Rhodobryo-Piceetum caricetosum pilosae; Rh-Pic abiet. sib - Rhodobryo-Piceetum abietetosum sibiricae; *Помечены неопубликованные материалы |
| Автор |
Место |
Синтаксон |
Размер пл., кв.м |
Деревья и кустарники |
Травы и кустарнички |
Мохо-образные |
| В.Н. Коротков |
Московская обл., Домодедовский р-н* |
Rh-Pic car.pil |
100 |
11 (6-15) |
19 (14-29) |
1 (0-4) |
| Л.Б. Заугольнова |
Московская обл., Серпуховский р-н* |
тот же |
25 |
6 (2-10) |
16 (7-28) |
нд |
| Л.Б. Заугольнова, И.И. Истомина |
Московская обл., Нарофоминский р-н* |
тот же |
25 |
6 (2-11) |
17 (8-27) |
6 (1-13) |
| А.И. Широков |
Нижегородская обл., Килемарский заказник* |
Rh-Pic abiet. sib |
100 |
9 (8-11) |
23 (15-28) |
нд |
| М.С. Хомутова, 1941 |
Нижегородская обл., Водораздел Неи и Какши |
тот же |
100 |
нд |
27 (19-34) |
4 (2-6) |
|
Экологическая характеристика |
| Характеристика местообитания |
| Тип ландшафта: Моренные,
водно-ледниковые и зандровые равнины. |
| Положение в ландшафте:
Приурочены к умеренно дренированным и увлажненным местообитаниям
водоразделов и склонов речных террас, оврагов и холмов. Положение
в ландшафте подчиняется правилу предварения Алехина: по
мере продвижения к югу эта группа перемещается с хорошо
дренированных склоновых позиций на более плоские, пониженные
и увлажненные, с некоторой задержкой дренажа (см. обзор
– Заугольнова, Морозова, 2004). |
| Почвы: От слабооподзоленных
и подзолистых с оглеением до бурых лесных
с разной мощностью гумусового горизонта; как суглинистые,
так и супесчаные. |
| Положение в экологическом
пространстве. Рисунок |
| Занимают центральную область
экологического пространства по основным факторам среды:
умеренного и слегка повышенного увлажнения, слабой кислотности,
высокой трофности почв. Эта область частично перекрывается
с областью ельников высокотравных и ельников нитрофильных. |
Сукцессионный статус |
Группа сложных ельников и широколиственно-еловых
лесов объединяет основные зональные сообщества и является
на равнинной территории одной из самых богатых во флористическом отношении и довольно разнообразной в типологическом отношении. Эти сообщества
можно рассматривать как довольно близкие к типу зонального
климакса для большей части территории южной тайги. В подтайге
эти сообщества тоже ранее занимали значительные территории.
Как ельники, так и елово-широколиственные леса этой группы
уничтожались в первую очередь, поскольку почвы обладали
высоким плодородием, а леса – продуктивностью (Ниценко,
1960).
Поэтому на водораздельных пространствах южной тайги и
подтайги большинство современных лесов этой группы
с абсолютным господством ели
являются производными сообществами, сформировавшимися
под влиянием многовековой лесохозяйственной деятельности
(выборочных рубок, пожаров, посадок леса), нередко чередовавшейся
еще и с агрикультурными мероприятиями. В результате степень
нарушенности/восстановленности получилась неодинаковой
у разных сообществ . Так, в разных регионах имеются, с
одной стороны, ельники, которые по составу травяного покрова
соответствуют «сложным», но практически не содержат широколиственных
деревьев и кустарников, а, с другой стороны, существуют
разнообразные варианты сообществ этой группы, различающиеся
по соотношению хвойных и широколиственных видов в древостое
и подросте. Если в северной части южной тайги незначительное
участие широколиственных деревьев в древостое еще можно
связывать с климатическими условиями, то в ее центральной
части и тем более в подтаежной зоне это можно рассматривать
только как свидетельство того, что неморально-бореальные
ельники являются производными сообществами от елово-широколиственных
лесов. Даже при кратковременных наблюдениях в сложных
ельниках отмечены тенденции ослабления позиций бореальных
видов и усиления неморальных. Так, для одного из вариантов
сложного ельника установлено, что на участках с разреженным
травяным покровом сначала разрастаются зеленые мхи и бореальное
мелкотравье, затем доминантом становится кислица, и далее
– зеленчук и осока волосистая (Речан и др., 1993 – стр.
68).
Авторы книги «Леса Северного Подмосковья» (Речан и др.,
1993), судя по использованию ими ссылок, ставят знак равенства
между ельниками кисличными Рузского района (Киселева,
1971) и ельником кисличным в Шолоховском лесопарке (Хлебниковская
вторичная моренная равнина – Речан и др., 1993). Однако
они не идентичны: первые являются производными от широколиственных лесов,
а вторые – от хвойно-широколиственных.
Поскольку в экологическом пространстве
область сложных ельников заметно перекрывается с областью
высокотравных ельников, более богатых по флористическому
составу, то при непосредственном соседстве тех и других
сообществ в ландшафте вполне вероятен такой тренд сукцессионного
развития сложных ельников как внедрение в них различных
видов лесной и опушечной флоры из высокотравных ельников,
т.е. повышение их флористического богатства. |
Созологический статус |
| Эта группа сообществ характеризуется
весьма сильным варьированием видового богатства в зависимости
от степени антропогенных нарушений. Наиболее сохранившиеся
сообщества с полидоминантным травяным ярусом заслуживают
внимания в качестве рефугиумов и потенциальных центров
дальнейшего восстановления зональной растительности. |
Характеристика ареала |
| Эта группа типов широко представлена в южной
тайге и подтайге и описана практически во всех областях:
Ленинградской, Новгородской, Псковской (Коротков, 1991;
Василевич, 2004), Нижегородской (Соколов, 1931; Широков,
1998), Тверской (Шапошников и др., 1988), Кировской (Смирнова,
1954), Московской, Смоленской, Костромской (Заугольнова,
Морозова, 2004; Смирнова и др., 2006). |
Источники информации |
| Василевич В.И. Елово-широколиственные леса Северо-Запада Европейской России// Бот. журн. Т. 89. № 8. 2004. С. 1249-1263. |
| Заугольнова Л.Б., Истомина И.И. Неопубликованные
описания. Московская обл., Наро-Фоминский р-н, д. Малинки.
[Сводная таблица
описаний] |
| Заугольнова Л.Б., Морозова О.В. Распространение
и классификация неморально-бореальных лесов // Восточноевропейские
леса: история в голоцене и современность. 2004. Книга
2. С. 13-62. Опубликованы сводные таблицы следующих описаний:
Московская обл. (Приокско-террасный заповедник), автор
– Л.Б. Заугольнова [Сводная
таблица описаний]; Нижегородская обл. (заказник «Килемарский»),
автор – А.И.Широков [Сводная
таблица описаний]; |
| Коротков К.О. Леса Валдая. М:
Наука, 1991.160 с. [Сводная
таблица описаний]. |
| Коротков К.О., Морозова О.В. Некоторые лесные
сообщества союза Сarpinion betuli в Подмосковье.
Депонир. в ВИНИТИ, № 3395-В88. М.,1988. 33 с. [Сводная
таблица описаний] |
|
| Соколов С.Я. Типы леса восточной части Баково-Варнавинского
учебного леспромхоза //Природа и хозяйство учебных леспромхозов
лесотехнической академии. Вып. 2. М.-Л.: Госсельхозгиз,
1931. С. 115-251. [Сводная
таблица описаний] |
| Хомутова М.С. Очерк растительности водораздела
рек Неи и Б. Какши //Уч. зап. МГПИ им. Ленина. 1941. Т.
30, вып. 1. С. 79-99. [Сводная
таблица описаний] |
| Шапошников Е.С., Коротков К.О., Минаева
Т.Ю. К синтаксономии еловых лесов Центрально-Лесного заповедника.
Ч.1. Неморальные и травяно-болотные ельники. Депонир.
в ВИНИТИ, № 4083-В88. М., 1988. 71 с. [Сводная
таблица описаний] |
| Дополнительный
список литературы (открыть
в отдельном окне) |
| |
| |
| Составитель: Л.Б. Заугольнова, О.В. Морозова |
|